第十讲
一、眼动对空间视觉的影响
二、几何图形的视错觉
三、触觉的空间知觉
四、意外事件引起的失明和先天性失明
五、为什么视觉的对象不颠倒?
六、关于空间构造理论的结论性评说
一
我们上述的一切考察都以经验的事实(empirical facts)为基础。反射运动定
律,肌肉的感受性,光觉和触觉中的部位差异(local differences),感官刺激的
长时暴露引起的衰竭(exhaustion)――所有这些都是可以用经验加以验证的现象。
但是,关于这些考察的结论,我们似乎把牢固的经验基础远远地置于脑后。我们根
据特定因素的联想协作(associative coopration)提出了一种空间的心理结构
(psychological construction of space)。是不是只有经验才能达到这一点呢?
难道空间不是生来就有的心理拥有(possession of mind)吗?或者,如果不是这
样的话,那么它至少不是我们知识中的全新要素吗?它是独特的(sui generis),
而非导源于其他任何东西吗?
空间知觉(perception of spece)是我们知识中的一种新要素(new element),
这是确定无疑的。但是,在这个意义上说,每种心理事实都是新的,因为它导源于
我们心理生活中一些要素的某种特定结合。这一结合的定律是,心理结果的特性是
无法预料的,原因在于进入到它们中间的要素特性,尽管我们以后能够见到这些要
素的联结及它们的结合。于是,在我们完成了复杂的推论过程以后,我们认识到从
这些前提必然地得出此项结论。但是,正如与这些前提作对照那样,仍然有一些新
的东西,一些必须通过明确的思维活动加以演绎的东西。认为空间知觉是天生的心
理拥有,或者认为空间特性是我们视觉和触觉的一种原始特征,这样的概括性断言
是不会有任何收获的。不仅这些陈述无法得到任何证明,而且那些系统阐述过它们
的陈述也不曾花过力气去考察摆在它们面前的心理学问题。作为一个问题,它肯定
存在着――以确定眼动定律和触觉器官的定律,以及与它们联系着的感觉联想(se
nsational associations)是否对这些感官知觉施加任何影响。
我们借助眼睛可以相当正确地比较出距离,这是为人们所熟知的事实。但是,
往往会发生这样的情况,两种并不精确相等的距离反而被认为是相等的,正像在简
单感觉的情形中那样,一种差别的知觉(perception of a difference)只有在达
到一定程度的量值(magnitude)时方才变得清楚起来,并在每种特定的情形里由所
涉及的感官特征所决定。在目前的例子中,就像在感觉强度(sensaion-intensit
y)的范畴中一样,我们能够通过测量来确定两种量值之间的差别达到多大才可恰好
被感知。
我们画两条长度相等的或几乎相等的水平线,然后询问一名观察者(他对两根
线的客观关系一无所知),这两条线在他看来是相等还是不相等。如果开始时我们
画的两条线长度相等,并将其中一条线的长度逐步延长,那么我们便将到达一个点,
在这点上那条逐步延长的线恰好能被观察者觉察到比另一条线更长。于是,实验暂
时中断,对两条线之间的长度差别进行度量。如果以各种长度反复进行这一过程,
我们便将获得一系列差值(different values),它们告诉我们对距离差别的感知
随着被比较的距离的逐步增加而变化。
这种实验与我们早些时候确定感觉依赖于刺激的实验基本上相同。我们仅仅用
空间量值替代了刺激量值而已。如果我们开始画的两条线的长度均为1分米,如果我
们逐渐增加其中一条线的长度,当增加的长度达到大约l/50分米,即2毫米时,两
条线的长度差别便可以看出来。但是,如果我们画的线开始时长度为1/2分米,那
么可以辨别的长度差别也相应减少。现在,确定这种差别为1%分米,或者1毫米,
不管我们用哪种测量标准,这种比例始终保持不变。在一定的上限和下限之间,这
种差别始终接近整个长度的1/50。在图18中,有两条水平线,左边一条长度为26毫
米,右边一条为25毫米。我们立即可以看出左边一条较长;不过,假如左边那条线
再截短一点,那么差别就觉察不到了。你可以借助实验来使自己相信,如果这些线
的长度延长2倍或3倍,那么它们的差别也一定会扩大2倍或3倍。
很明显,我们先前曾发现一条定律,即恰好能被觉察到的感觉差异有赖于刺激
差异,现在这条定律同样适应这里的情况。“恰好能被觉察到的空间距离的增加与
整个距离始终保持同样的比例”。显然,这种巧合可以十分简单地用下述事实来解
释,也就是说,我们在感觉方面拥有对空间关系知觉的测量,为我们直接提供这种
测量的感觉是那些来自眼球运动的感觉,感觉的强度肯定随眼睛注视路线的长度而
增加。
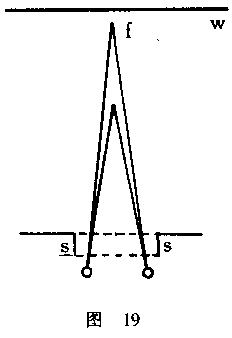
在我们面前有一只匣子ss(见图19),一面打开,另一面有一水平的细长裂缝,
通过该匣子,两眼能看见白色屏幕w,而不是看见房间里面的任何其他物体。现在,
我们在屏幕和眼睛之间悬一根垂线f,用重物将垂线拉紧。每只眼睛本身将采取这样
的位置,即使垂线f在黄斑上形成一个映像,这是视觉最清晰的地方。从这一点穿过
眼的中央所经空间的连线称为视轴(visual axis)。因此,我们可以说,两只眼睛
的视轴相交于f点上。如果我们现在改变一下垂线的位置,即把它挪近些或者距离眼
睛远些,结果视轴相交所形成的角度也同时发生变化;因为眼睛始终追随着那条线,
并一直指向着它。如果垂线移向更远的距离,那么两眼便会向外,视轴的相交角变
得更尖锐;如果垂线移动得更接近眼睛,那么眼睛便会向内,相交角也就更钝。当
我们了解了垂线距离的这种变化时,我们便可以容易地确定每只眼睛围绕其中央转
动有多远。如果垂线一点一点地移动,那么它的距离变化将难以被感知到;也就是
说,眼睛围绕其中央的转动如此微小,以至于相伴的运动感觉无法被觉察。只有当
垂线位置的改变已经达到一定的量值时,运动感觉方能被觉察,我们才能感知到这
条线已被移近还是移远。这种界点的确定必须经过长期的系列实验,并且以垂线离
眼睛的不同距离来进行实验。我们应当发现,当两个视轴实际上平行时,也就是说,
当两眼近似于处在休息位置时,眼睛就其本身的运动来说便拥有最佳的感受性。在
这样的情况下,如果每只眼睛围绕其中央的转动只达到一度的1/60(角度1'),我
们便可感知距离的变化。
但是,一俟眼睛内转到相当大的距离――当然,这种情况发生在垂线向更近处
移动时――则恰好能被觉察到的运动就会变得很大。我们将会发现,这种恰好能被
觉察到的运动的量值是按眼睛距离它休息时的位置成正比增加的。
很显然,这里的讨论仅仅是进一步确定恰好能被觉察到的感觉有赖于刺激的普
遍定律。眼睛的内向转动产生了明确的运动感觉。这种运动的量值与刺激强度相对
应;业已出现的运动越大――也就是说,已经在运作的刺激越大――运动的增加或
刺激的增加也肯定越大。如果对运动感觉的感知与外部感官的感觉一样,遵循着同
样的定律,那么便可以预期,与相等的能够觉察到的感觉的增加相对应的运动的增
加将始终与已经存在的整个运动成正比。事实上,实验证明,这种关系差不多是不
变的。甚至与这个规律相应的偏离,我们已经发现在外部感官感觉的情形中也是站
得住脚的。也就是说,当运动的程度很大时,分辨的精细度比之我们根据该定律所
作的预期要小一些。但是,运动的增加恰好足以产生能够觉察到的感觉,差不多达
到整个运动量值的1/50。这一结果与我们从空间量值的比较中业已获得的结果完全
一致:当一条较长的直线与一条较短的直线之间的差别达到后者长度的1/50时,较
长的线便恰好能与较短的线区分开来。但是,如果空间距离的知觉与眼睛在扫视这
段距离时所作的运动努力成正比,那么,我们便必须得出结论,运动的努力是知觉
的标准。由于我们只能通过运动感觉方可具有努力的知识,因此后者的影响也得到
证实。
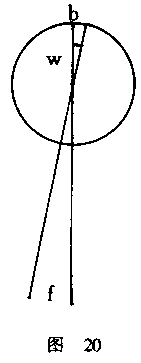
关于运动感觉与距离估计相联结的实验可由下述观察予以补充。我们并排地悬
吊两根黑线,黑线后面的背景是明亮的,黑线与背景之间保持一定距离,要求用一
只眼睛凝视这两根黑线(图对)。然后,我们逐渐离开这两根黑线,一边移动一边
不断地注视着这两根线。由于距离远的物体看起来比近的物体小,因此两根线之间
的距离在不断缩小,直到达到某一点,在那里两根线差不多合二为一。我们可以理
解,当我们离开物体时,物体的尺寸变小,这是由于物体在视网膜上的映像变小的
缘故。因此,实验表明,视网膜映像的两点具有一定的量值,在这个量值之下,就
无法感觉到它们是分开的了。这种视网膜映像(b)的量值,或者说相应视角(w)
的量值,是可以确定的,因为两线之间的距离和它们距离眼睛的远近都是已知的。
我们发现,当两个视网膜映像之间的距离变得如此之小,以至于眼睛只需要转动1分
(l'),便可先把第一根线然后再把另一根线带到视网膜的同一点上时,两种映像
就合而为一了。但是,正如我们上面发现的那样,与恰好能觉察到的眼球运动的量
值具有同样的量值。因此,接下来的问题是,处于休息状态的眼睛以同样程度的正
确性感知空间中物体的距离,也就是说,以眼睛在最有利的条件下感知自身运动的
正确程度去感知空间中物体的距离。这里所谓的最有利条件,是指运动以视轴平行
为开端。眼睛在认识空间距离时能够达到的限度与它对自己运动感觉的感知限度相
一致。
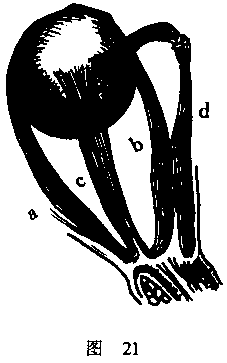
空间感觉对运动感觉的依赖(对此,我们已经根据基本的实验进行过推论)为
其他许多视觉现象所进一步证实。眼睛的肌肉总的说来是对称排列的(symmetrica
lly arranged)。因此一种肌肉(a),即外直肌(rectus externus),使眼睛向
外转动,而另一种肌肉(b),即内直肌(rectus internus),则使眼睛向内转动
(图21)。这两种肌肉在维度(dimensions)上差异只不过一点点,两者处于一个
水平面,该水平面穿过眼球中央。所以,它们的位置对它们产生运动的可能性极为
有利。这种完全相似的条件使得下述情况很明显,也就是说,由相等强度的旋转引
起的运动感觉将具有差不多相等的强度,而不论这些旋转是向内还是向外。我们在
向上运动和向下运动方面发现同样的情况。眼睛通过单一的肌肉(c),也就是上直
肌(rectus superior)而向上转动,上直肌在眼窝内的上部稍向前倾,而且附着于
眼球的上部,在中间稍稍向外。它的活动得益于另一种肌肉的运作,这种肌肉在我
们的图示里被眼球遮住了。这种肌肉称为斜下肌(obiquus inferior),位于眼窝
的下部,从前面向后向外,联结着眼球的后表面。这些肌肉在排列上同样是对称的,
通过这些肌肉,向下运动得以实现。与肌肉c相对的肌肉位于眼球的下方一侧,称为
下直肌(rectus inferior),其运作得到d,即斜上肌(obliquus superior)的帮
助,斜上肌是向前和向内运动的,并牵拉眼球的上表面。由于这些肌肉是对称分布
的,因此运动的努力与我们不论把眼睛向上翻或者向下翻所作的努力是同样的。另
一方面,在使眼睛向外转或者向内转的肌肉排列和使眼睛向上翻或向下翻的肌肉排
列之间存在相当大的差别。如果在这一关系中也要求相似性的话,那么这些肌肉必
须这样来放置,也即上直肌卜)(它使眼睛向上转动)和下直肌(它在眼球的另一
边,使眼睛向下)应当附着于一点,以便最有效地促进它们将实施的运动。然而,
正如我们的图示表明的那样,实际情况并不这样。C的方向比a和b的方向更为倾斜。
因此,花费同样的力气,前一肌肉在使眼睛向上转动的距离要比后一对肌肉中的任
何一条使眼睛向内或向外转动的距离稍稍少一点。鉴于这一原因,它得到了第二种
肌肉的帮助。所以,产生上下运动所需的用力程度一般说来要比产生向外或向内的
相等伸展的运动更大一些,据此,运动感觉也更强烈;而且,我们必须期望发现,
垂直方向的距离将比水平方向的同样距离显得更长一些。作为事实,这是无疑的。
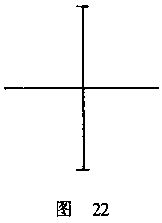
如果我们用相等的臂长画一个十字,那么在垂直方向上将显得长一些(图22);而
在其他一些图形中,如在正方形或矩形中,垂直距离同样会被估计过高。
二
在估计垂直距离和水平距离方面出现的这些差别是十分重要的,不过,它们并
不是用眼睛进行测量时发生的唯一错误。一个类似性质的较小差别可以在一根垂直
线的上半部和下半部之间观察到,也可以在一根水平线的内部和外部之间观察到。
因此,严格地说,在十字形(图22)的四个臂长中,没有哪一个会看上去与其他任
何一个恰恰相等。这些较小的差别在每个例子中也与肌肉的不对称排列相一致。当
我们在上面讲到两种肌肉a和b(它们使眼睛向外和向内转动)在其维度方面只有极
小的差别时,当然意味着它们并不完全相似。实际上,b(内直肌)要比得到更强的
发展,也许是由于视轴的辐合运动(converging movements)在所有例子中均占支
配地位。我们乐于从事唾手可得的事情――例如,我们正在凝视邻近的物体,于是
b比a得到更多的练习。还可以相应地观察到,一根二等分的水平线的外面一半要比
里面一半显得更长;较弱的肌肉需要用更大的力去产生相似的运动,而更大的用力
则与更强的肌肉感觉相伴随。为了实现这种明显的不相等,当然必须闭起一只眼睛。
对右眼来说是外侧的东西,对左眼来说则是内侧;双目视觉破坏了这种不相等。还
有一种类似的差别,这种差别在双目视觉中也不会消失,它就是视野上半部和下半
部之间的差别。如果我们仔细看图22中的十字形,我们可以看到,垂直线的上半部
显得比下半部长一些。这种差别也是与肌肉分布的不对称相一致的。那些把眼睛向
下拉的肌肉比起那些使眼睛向上翻的肌肉得到更有力的发展――也许由于同一原因,
我们发现在内直肌和外直肌的例子中也是如此。由于视轴通常有点指向下方――当
我们凝视近距离物体时,这一点尤其确切――使眼睛朝水平线下面转动的肌肉得到
更多的练习,从而使一种向上运动要比具有相等伸展的向下运动花费更多力气。
眼球肌肉不对称分布的视觉效果使我们能够正确预测距离估计方面的其他一些
不正常情况,这些情况在实验中得到证实。你们都知道,如果我们用小步子走一段
距离将比我们用大步子走一段距离更加感到疲劳。眼睛同样适用这种情况。通过一
段没有阻断的途径要比通过一段距离相等却常被阻断的途径,前者花的力气较小。
如果我们将一根直线分成相等的两段,然后将其中一半分成无数更小的段,那么细
分的部分要比未经细分的部分显得更长一些。当然,这种实验可以各种形式发生变
化。例如,一只细分的角度要比一只未经细分的角度同样显得更大些;一个平面图
形,当被分成无数小块时,要比面积虽然相等但未分成小块的平面图形显得更大些,
如此等等。这些现象(可以在几何图形中得到充分的观察)称作“几何图形的视错
觉”(geometrical optical illusions)。它们都是眼动感觉在空间视觉活动中协
同作用的令人信服的证据。
触觉的空间知觉在许多方面不同于视觉的空间知觉。这种差别可能部分地由于
下述事实,即在我们心理生活的正常发展中,比起皮肤来,眼睛是一种发展得更加
完善的工具,眼睛的特定发展看来先于精细的触觉的特定发展。当然,这并不意味
着两种过程是截然分开的;恰恰相反,它们是相互交叉的,每一种过程影响和帮助
另一种过程。但是,至少对于人类和高级动物来说,视觉是一种较早的活动;触觉
恰恰是受视觉引导和指导的,而不是相反。
如果我们还记得皮肤的压觉(pressure-sensations)始终受视觉的影响,那么
我们将会发现,依附于皮肤压觉的部位关系(loeal reations)首先是视觉的。但
是,触觉属于这样一种性质,它能够在一定程度上摆脱这种影响。对皮肤来说,正
像对眼睛来说一样,感觉的特定性质有赖于印象的定位(locality),并随地点的
变化而变化。因此,这种定位最终会被感觉本身的部位色彩所认出,而毋须召唤眼
睛来协助抉择。当眼睛一旦确定了定位与其部位色彩的关系,我们便能够将确定的
感觉归入皮肤感受表面的正确位置上去。
由此引申出去,我们可以发现,印象的空间分辨将不再依靠运动,或者依靠与
此相伴的感觉的生动性和可比性,而是单单依靠感觉的部位色彩中或大或小的差别。
如果皮肤的两个相邻部分在这方面的差别极小,那么我们便无法区分由这两部分产
生的感觉。当它们对感觉特性确实不同的皮肤的两个相邻部分产生影响时,我们将
只能感受到这些印象在空间上是有差异的。很显然,这种限度不是固定的和不变的,
通过密切注意我们的感觉,我们将能够在彼此接近的两个印象之间作出区分。以此
方式,我们在实验中观察到的练习之影响便有了自然的解释。
我们发现了存在于皮肤表面各个点上的分辨能力差异,这些差异将以同样方式
有赖于一种精细程度,而部位感觉差异则是根据这种精细性来分级的。这种差异实
际上是很大的。例如,用圆规的两只脚(两脚之间的距离甚至小到只有1毫米),在
指尖上我们能清晰地区分出两种独立的印象;可是在背部皮肤上,我们区分两种独
立印象的距离就必须达到60毫米。因此,可以把整个皮肤视作是感受点的分级系统。
但是,这些感受点并不是以感受性的顺序均匀地排列在皮肤上面的,而是彼此之间
具有不同的距离,并不同地分布着。除了这些肤觉(cutaneous sensations)的自
然特性以外,眼睛对它们的控制可能也影响这种分级。在皮肤上,并非所有部分都
相等地隶属于视觉的控制;许多部分,像背部的皮肤,完全处于视线以外;皮肤的
其他部分,例如手和手指,则特别受到眼睛的控制。还必须记住,皮肤上的所有部
分并非自然地得到同样的练习。两只手,尤其是指尖和手指,则经常得到练习的机
会;次于两只手的是嘴唇和舌头。由于这种练习量的自然差别,皮肤的空间分辨能
力的进一步发展(这是我们通过有意练习而达到的)在皮肤的不同部位有很大差别。
例如,在指尖,这种空间分辨能力的进一步发展就较小,而在手臂的上部和下部,
其发展就令人注目地大;在几小时内,这种分辨能力可以增加1倍或者4倍。事实上,
练习的好处很快就会消失:经过24小时以后,它的感受性便减弱;过了几星期或几
个月以后,这种感知性完全消失。但是,这种结果并不限于直接练习的那部分皮肤。
例如,如果右手手背的感觉精细度翻了1倍,那么,左手手背的感受性也会等量增加,
尽管左手尚未得到过练习。同样的结果可以从皮肤的所有对称部分中获得。与此同
时,练习的效应不会扩展到这些对称部分之外的地方去。例如,练习右下臂或右脸
颊,可能同时有助于练习左下臂或左脸颊;但是,对上臂、胸部或前额不会有任何
的作用。这种特殊的结果必须根据练习中涉及的心理过程来解释。在练习中,我们
学会了注意感觉差异,这些感觉差异在我们练习之前是注意不到的。现在,皮肤对
称部分感觉的部位特征是十分相似的。因此,如果我们学会了注意一侧的较小感觉
差异,我们也将学会对另一侧相应部位的较小感觉差异予以注意。特别是左侧和右
侧,在分级的精细度方面具有完全的对应,而且在部位色彩从一点到另一点的变化
速度方面也完全对应。当然,当我们处理不对称的部位时,情况便不同了;感觉及
其分级如此不同,以至于在一个部位得到的经验不可能应用于另一部位。或者,这
种先前的经验至多只有一些价值,因为一般说来注意由于练习而变得更加敏锐了。
我们已经看到,在皮肤上刚刚能够觉察的差异也许不是由运动感觉决定的(这
是就正常的个体而言),而仅仅是部位感觉差异的分辨结果。与之相同,我们对于
触觉印象之间空间距离的增加或减少的判断,仅仅有赖于我们根据每一种感觉的部
位色彩而对每一种印象的位置的了解,或者,更正确地说,有赖于把两者联系起来
的持久联想。但是,这种了解是依靠视觉的帮助而获得的。我们根据受刺激部位的
记忆意象而对皮肤上的距离作出是长还是短的判断,这种记忆意象是受刺激部位的
感觉引起的。这种记忆意象并不受制于通过这段距离时所需的运动;它仅仅受制于
由视觉帮助我们形成的观念,即皮肤上的每个部位是由它的特定感觉特性决定的。
一种明显的推论是,空间距离的分辨,不论是大是小,只要皮肤表面的感受性保持
不变,这种分辨也保持不变。这是一种我们通过实验而实际获得的结果。如果11毫
米的一段距离恰好能被觉察到与10毫米的一段距离不同,那么,我们也能对20毫米
和21毫米进行区别,对30毫米和31毫米进行区别等等。总之,对于皮肤感受性而言,
不是相对的(relative)恰好能被觉察到的距离差别保持不变,而是绝对的(abso
lute)恰好能被觉察到的距离差别保持不变。这一规律的例外可以下述事实来解释,
即在长距离中,我们对相邻皮肤点的分辨精细度有相当大的变化。
四
触觉的正常发展比视觉的正常发展来得更晚;结果,触觉用于空间的测量是从
视觉中获得的,而不是从四肢的动觉中获得的。由于这一原因,触觉反射的机制和
触觉发展的定律将不具有巨大的重要性,这种重要性是在视觉发展中依附于它们上
面的。它们的影响必然降低到这样的程度,也就是说,它们的影响由于视觉的支配
作用而遭破坏。但是,这种破坏仅仅是部分的。阻碍视觉影响的每一种影响增加了
触觉的独立性,并有助于使它发展到这样一种程度,也即在一般条件下不可能达到
的一种程度。意外事件引起的失明向我们表明了这方面引人注目的变化:肌肉变得
更加容易反应;最小的触觉刺激引发一些运动,这些运动使外部物体与皮肤的不同
部位相接触,尤其是与最敏感的部位相接触。在一些罕见的例子中,即在心理发展
的初期便不存在视觉的支配性影响的场合――也就是在先天性失明的例子中,触觉
运动所起的作用仍然很大。
先天性失明的人被迫从触觉中建立起他们的全部空间世界。他们以非凡的完整
性做到了这一点。在正常生活中处于低级发展水平上的那种官能达到了完美的程度,
在分辨的精细度方面至少可以与间接视觉(indirect vision)(也就是视网膜边缘
部分的视觉)的分辨精细度相比较。只有在一个方面,皮肤才比眼睛略逊一筹:它
要求与它的印象直接接触。
那么,先天性盲人是如何获得空间距离的观念或空间延伸的物体的观念的呢?
他们具有来自皮肤的压觉,以及来自正在探索的四肢的运动感觉。根据这些,他们
建立了空间知觉。这种构建的手段显然可以在两种感觉系列的联想中被发现(正如
在视觉中一样),而联想则是由反射机制的一致作用形成的。但是,后者在盲人中
肯定要比亮眼人需要更完整的发展。因为,每一肢体被带入与皮肤某个明确部分的
反射联结中去,感觉的部位差异最终与运动的明确感觉相联系。因此,在皮肤的每
一个区域存在着某个中心点(尽管这个中心点可能会变化),所有邻近的感觉都被
归之于该中心点。于是,皮肤的各独立部分被带入到彼此的关系中去。而且,通过
原先不同的感觉系统的相互联结,整个肤觉组合成一个单一的系统。不论何时,只
要分开的四肢开始彼此接触,这种相互联结就必然倾向于受到影响。以此方式,便
能对彼此独立的触觉器官及其感觉中心之间的距离进行某种测量,尽管是不完全的。
毫无疑问,这一发展过程将需要比视觉的训练花更长的时间。后者用单一的活
动便可完成;但是需要大量的连续活动,它们联合成一种共同影响的能力是由于以
下事实,即它们都属于类似的性质。也就是说,引起视觉的空间知觉的过程必须为
触觉而多次重复。现在,就像我们通常用黄斑注视任何事物以便获得清楚的感知那
样,盲人也被迫通过皮肤各部分感受性的巨大差别来使这些部分专门用于精细分辨
方面的感知。具有这种能力的触觉器官是双手。盲人经常用双手练习触觉,甚至更
多地在运动中加以练习。当然,仅仅用触觉显然还不足以确切地感知空间关系,原
因在于,如果一个物体的各部分并不恰好位于同一平面上,则皮肤的压觉不可能为
它们提供任何反映。于是,双手细微的触觉运动,尤其是手指的触觉运动,在盲人
中间是异常活跃的,因而具有十分巨大的重要性。通过他们的触觉,物体的空间特
性便能确切地被感知,这里,部分地通过与触觉器官各部分的连续接触,它们能进
行最精细的分辨,部分地由于压觉和动觉的连续联结。但是,我们经常发现,盲人
不能像我们用视觉获得最复杂图像的确切观念的那种速度去感知甚至十分简单的空
间关系。他们的触觉和动觉必须根据该复杂图像的各个部分逐步构建那个物体。
五
由此可见,事故性失明者和先天性失明者建立空间知觉的缓慢而又不完整的发
展过程进一步确定了我们的假设,即一般说来,视觉胜过触觉。在坚持这个观点时,
我们与旧心理学中相当流行的观点发生了冲突,这种观点目前尚未完全被抛弃――
该观点认为,视觉更可能受制于触觉的训练。据称,我们用手触摸的东西,比起在
很长距离外对我们影响的东西,在感知上更为确定。但是这种观点恰恰忘记了,相
似的两个物体都使感觉神经产生一种印象,而在缺乏相关心理过程的情况下,这些
东西在印象的起源上便无话可说了。
但是,也有一种特殊情况似乎支持这一观点,即触觉对视觉的发展来说是必要
的。我们看到的物体处于它们的自然位置中,而不是颠倒状态。但是,外部物体在
视网膜上产生的映像却是颠倒的。眼睛是一种光学仪器,它由一系列曲面(curved
surfaces)组成,曲面在视网膜上投下一个位于视野内的各种物体的微型映象。然
而,这一映像的空间关系恰恰是物体本身空间关系的颠倒:如果物体用脚站立,那
么视网膜映像就用头站立,反之亦然。因此,只要假设视觉活动以形成视网膜映像
而告结束,那么我们关于物体正确向上的视觉必然成为一种谬论。但是,心理对视
网膜映像的了解究竟是什么呢?我们仅仅了解了其存在及其颠倒的位置,正像物理
学家和生理学家关于物体的了解一样。为了能够像这些映像的实际情况那样去感知
它,我们必须假设在视网膜后面还有另一只眼睛。而且事实上,这种假设不时地被
认为是一种可能的假设。当然,第二只眼睛不可能真正存在;可是,据假设,当映
像影响头脑时,它再次被头脑所颠倒过来,正像它被第二只眼睛颠倒过来那样――
一位机灵的哲学家说得好,不将这种永恒的颠倒状态归之于头脑,而让它自己倒立,
反而会简单得多,这样一来它的颠倒可能会使视网膜上反映的颠倒世界拨正过来。
根据我们自己关于空间视觉调查的观点,这种困难能够十分简单地得到解决。
它只是一系列作用于我们头脑的视网膜映像的部位色彩感觉。只有通过眼睛的动觉,
头脑才能学会将这些东西联结成一种空间顺序。但是,动觉告诉我们关于物体位置
的哪些东西呢?随着眼睛的转动,它从外部物体的一点移向另一点。在围绕着它的
中心从上到下的移动过程中,它从上到下传递一个物体。它将视网膜映像的各个部
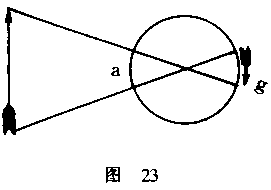
分依次地带入最清晰的视觉点上。现在,当眼球的可视部分a向下移动时,眼球背后
的黄斑g将向上转动(图23);随着前面的a点凝视着物体的不同部分,g点也在以同
样精确的方式详细审视着视网膜的映像。因此,如果物体在空间中的位置是根据运
动推算出来的话,那么,视网膜映像就一定是倒置的,因为只有这样的情况才有可
能使运动与物体的实际位置相一致。倒置的视网膜映像对视觉来说是十分必要的,
而远远不是一种谬论。视网膜映像肯定会被颠倒过来,即使眼睛中光的折射定律没
有使这种颠倒成为生理上的必需。
当然,进一步的问题也可能会被提出来,例如我们如何知道我们在使眼睛向上
或向下转动。难道“上”和“下”不是关系的概念吗?难道它们不是正在感知的被
试和他的空间位置的先决条件吗?事实上,正是因为上和下是有关系的,我们才能
将顺序引入空间视觉的世界。如果我们具有上和下的绝对方向的知觉,我们便应当
认为,不论白天还是黑夜,只要情形许可,我们便可以头着地倒立;那是就地球的
转动而言的。我们之所以不这样认为的原因是,由于我们将自己视作一切空间关系
的中心。上和下,像右和左一样,都是一些只有涉及我们自身时才具有意义的术语。
在我们的空间知觉中,当区别上和下的时候,我们不断地以我们自己的身体为参照:
当任何物体与双脚处于同一方向时,我们称之为“下”;当任何物体与头处于同一
方向时,我们便称之为“上”。
但是,仍然存在一种异议,这种异议似乎反对运动对空间感知的影响,对此我
们已经认识并确实证明在许多情形里存在这种情况。人们可能要问,当我们从空间
角度看事物时,我们是否真的始终转动眼睛?我们是否必须实际地上下转动眼球以
便知道什么东西在上和什么东西在下?并非如此。我们用不到转动自己的眼睛,便
能感知事物的空间延伸,并将其空间位置一一排列。我们将如何对待这一异议?我
们可以像平时所做的那样,指出眼球运动的巨大速度,而我们最终却没有能力去观
察它们。可以这样假设,尽管我们认为眼睛处于休息状态,但它实际上进行着十分
快速的运动。不过,我们无法用此方式来避开困难:肌肉运动的速度不会像我们在
这个假设中想象的那么快。另一方面,我们可以用实验方法大大减少一种光的印象
的持续时间,以至于完全排除眼睛运作时眼球运动的可能性,例如,通过用电火花
的即时照明。在这种情况下,物体仍然会在空间上被看到。因此,毫无疑问,对于
每一种单一的空间知觉而言,运动不是必不可少的。
但是,还有一点不可不注意。我们必须经常将心理过程(mental processes)
与心理产物(mental products)相区别。后者可能有赖于先前发展过程中获得的一
种能力。在形成我们的空间知觉中,最初存在着一个因素,它运作着,以便空间知
觉的进一步完善,将此视作一切视觉的持久的和不可避免的条件,这是不必要的。
小孩子通过母亲的指导去跨出第一步,借此学习单独行走。那么,为什么不该也有
一些视觉条件,它们是单独运作的,或者至少是以它为主运作的,在视觉发展的第
一阶段便存在呢?
实际上,我们已经发现了这种视觉条件。视网膜感受点的相对位置是由一系列
集中分级的运动感觉决定的,并处于与相关的、部位色彩的光觉进行联想性联结的
状态之中。如果曾经体验过的印象第二次被提供,那么这些点就能通过它们的部位
色彩而被认出。因此,如果两种印象作用于两个视网膜点(后者在以前的场合因为
明确强度的运动感觉而被彼此分离),那么,经过对整个过程的频繁重复以后,我
们就能够将它们进行区别,而无需实际运动的发生及其伴随的感觉。当部位感觉差
异一俟从将它们分离的距离测量的运动感觉中获得,它们便保持这种测量,而不受
其起源的制约。一种明确的地点参照依附于部位色彩,在它后面,作为感觉性质的
真正特征完全消失。我们想象我们直接感知一种印象的定位,而实际上我们只感知
到感觉的特性,根据特性认识定位。当我们通过练习扩展了我们的空间分辨能力时,
我们认为我们感知空间差异的能力已经直接增强了,而事实上只有分辨小的感觉差
异的能力得到了提高。在这方面,对于视觉来说是正确的东西对触觉也适用,只是
后者(甚至当它已经达到了不同寻常的高度发展时)处于经常需要来自运动感觉的
进一步帮助的状态之中,这是由于它的部位感觉属性的不明确特征所致。因此“感
受”(sense of feeling)这个名称(有时用于触觉)是颇为重要的。原先我们用
眼睛“感受”物体,正如我们用手“感受”一样。但是,手仅仅是一种“感受”器
官,不仅因为它必须实际上与它感知的物体相接触(视网膜映象通过~段距离之外
的光的活动而产生),而且还因为它在接触以后,还要继续去“感受”;只有通过
两种感觉的结合,也就是压力和运动的结合,方能获得一种完整的知觉。
六
我已经用这样一种方式尝试性地描述了空间感知的现象,即将解释和协调这些
现象的理论作为其本身的结果。我向你们提供的这种理论是由一些事实直接提示的,
而且并未试图超越这些事实以外。但是,在我们考虑结论之时,我们不应忽略这样
的陈述,生理学家和心理学家仍然认为,他们可以完全省却对我们视觉和触觉进行
排列的任何一类解释,或者,至少可以认为,我们在上面讨论的这些因素之一足以
解释了一切事实。据假设,在前面的例子中――正如在旧生理学中的情况那样――
我们的观念的空间排列是在视网膜映象的各部分排列中被直接提供的;或者,正如
今天由于较慢的了解所陈述的那样,尽管在表述的形式上没有任何真正的改进,我
们在这里讨论的两种感觉中的每一种感觉从一开始便具有某种空间性质。现在,我
们无法否认这将是最方便的假设了。但是,同样无法否认的是,我们还完全不能解
释对我们理解和估计空间量值具有决定性影响的一切因素。在人们试图根据这一观
点对讨论中的因素进行适当处理的地方,已经发现有必要建立若干人为的和复杂的
次级假设,其中有的假设甚至走上了自相矛盾的极端。也许,这些情况对逻辑学家
具有某种价值,作为这些假设如何不该被构筑的告诫性例子,但是它们对心理学家
绝对无用。
当试图根据决定知觉活动的影响之一来为空间构造提供一种理论解释时,情况
就不同了。人们通常认为有可能单单按照运动和动觉来建立一种理论,或者完全忽
视视网膜和皮肤的部位感觉性质,或者认为它们完全不受运动的支配而起作用,并
且,像后者一样,其本身便足以适当解释这些事实了。在这些观点中,第一种观点
不可避免地导致这样的结论,即肌肉感觉具有特定的空间性质;后者将这种性质单
单归之于视网膜的感觉,或归之于这些感觉系列中的两种感觉。因此,它间接地意
味着回归到这样的观点,即外延的观念一般说来既不要求也无法作出任何心理学解
释。但是,仅仅承认印象的空间排列一方面受制于运动,另一方面由附属于静止的
感官特性以及与刺激部位联结的特性来决定,这是不够的。经验表明,这两种影响
如此密切地联结着,以至于离开了对方任何一种影响都无法起作用。关于这方面的
主要证据存在于下述事实之中,只能按照眼动定律来解释的那些影响在眼睛处于静
止状态时仍然保持着;对此,可以比较用电火花产生瞬间照明的例子。当运动被阻
时,上述关于水平线和垂直线相对长度的错觉以及其他一些类似的现象并没有消失,
尽管它们有时可能并不那么引人注目。
如果我们按照陆宰(lotze)的说法,我们把可能对空间观念的活动产生影响的
每一种感觉成分称作部位记号(local sign),那么,认为空间知觉由心理过程产
生的那些理论(既非优先,也不导源于特定的感觉性质),便可能被区分为简单的
部位记号理论和复杂的部位记号理论。第一种理论假设了动觉的部位记号,或皮肤
的部位记号,或两者兼有,然而在后者的情况下不允许两者的相互作用。另一方面,
复杂的部位记号理论则把外延的观念视作动觉的集中分级的部位记号的心理结果,
以及感官表面质量分级的部位记号的心理结果。空间知觉有赖于这两种感觉系列的
联结,尽管其中之一的一些成分只需再现便能起作用。这对强度系列(intensive
Series)尤为适用,它们与质量系列如此紧密地联结,以至于每一对明显不同的部
位记号将始终与动觉相联系,后者与它们之间器官通道的距离相对应。
亦凡公益图书馆(Shuku.net)扫校
下一讲 回目录